Powstawanie gatunków
Speciation jest o tym, jak tworzą się gatunki. Jest to główna część biologii ewolucyjnej. Darwin uważał, że większość gatunków powstała bezpośrednio z wcześniej istniejących gatunków. Nazywa się to anagenezą: gatunki przez zmianę, lub "ewolucją…
Speciation jest o tym, jak tworzą się gatunki. Jest to główna część biologii ewolucyjnej.
Darwin uważał, że większość gatunków powstała bezpośrednio z wcześniej istniejących gatunków. Nazywa się to anagenezą: gatunki przez zmianę, lub "ewolucją filogenetyczną". Przez większą część XX wieku sądziliśmy, że większość gatunków powstała w wyniku wcześniejszego podziału gatunków: kladogenezy. Powszechnie uważano, że większość podziałów gatunkowych jest spowodowana lub wspomagana przez mechanizmy izolujące.
Bez wątpienia fizyczna separacja gatunków, które kiedyś żyły razem, jest głównym czynnikiem. Ilustruje to wiele przykładów, z których niektóre omówione są poniżej.
Jednakże prace prowadzone w ciągu ostatnich 20 lat wskazują na inne przyczyny. Analiza sekwencji DNA istot żywych wykazała, że często dochodzi do hybrydyzacji pomiędzy spokrewnionymi gatunkami. Oznacza to, że geny zostały przeniesione w wyniku tych krzyżówek. To z kolei oznacza, że izolacja reprodukcyjna nie jest jedyną definicją gatunku, a specjacja nie zawsze wymaga allopatrii (gatunki muszą być rozdzielone reprodukcyjnie). Poniższe fragmenty ilustrują ideę, że fizyczna separacja miała pierwszorzędne znaczenie w powstawaniu nowych gatunków.
Galeria obrazów
10 Obrazy









Mechanizmy izolujące
Mechanizmy izolacyjne to rzeczy, które uniemożliwiają udane rozmnażanie między grupami w obrębie gatunku. Ustanawia się izolację reprodukcyjną populacji. Jest to szczególnie ważne dla koncepcji gatunków biologicznych, ponieważ gatunki są definiowane przez izolację reprodukcyjną.
Mechanizmy izolujące można podzielić na dwie grupy: przed i po zapłodnieniu.
Przed nawożeniem
Czynniki, które uniemożliwiają osobnikom kojarzenie się.
- Izolacja geograficzna: Gatunki występują na różnych obszarach i często są oddzielone barierami.
- Izolacja czasowa: Osobniki nie łączą się w pary, ponieważ są aktywne w różnym czasie. Mogą to być różne pory dnia lub różne pory roku. Okresy godowe poszczególnych gatunków mogą się nie pokrywać. Osobniki nie spotykają się ze sobą podczas okresów godowych lub w ogóle.
- Izolacja ekologiczna: Osobniki łączą się w pary tylko w preferowanym przez siebie siedlisku. Nie spotykają osobników innych gatunków o odmiennych preferencjach ekologicznych.
- Izolacja behawioralna: Osobniki różnych gatunków mogą się spotkać, ale nie rozpoznają żadnych wskazówek seksualnych, które mogą być podane. W większości przypadków osobnik wybiera członka swojego gatunku.
- Izolacja mechaniczna: Można próbować kopulacji, ale nie dochodzi do transferu spermy. Osobniki mogą być niezgodne ze względu na rozmiar lub morfologię.
- Niezgodność gametyczna: Następuje transfer plemników, ale jajeczko nie zostaje zapłodnione.
Po nawożeniu
Czynniki, które uniemożliwiają udane kojarzenie, takie jak niezgodność genetyczna, nieudatność mieszańca lub sterylność.
- Śmiertelność zygotyczna: Jajo zostaje zapłodnione, ale zygota nie rozwija się.
- Niezdolność hybrydy do życia: Zarodek hybrydowy tworzy się, ale nie jest zdolny do życia.
- Sterylność mieszańca: Mieszaniec jest zdolny do życia, ale powstałe z niego osobniki dorosłe są sterylne.
- Rozpad mieszańca: Mieszańce pierwszego pokolenia (F1) są zdolne do życia i płodne, ale dalsze pokolenia mieszańców (F2 i krzyżówki wsteczne) są niezdolne do życia lub sterylne.

Izolacja geograficzna
Uważa się, że jest to najbardziej powszechna przyczyna specjacji. Pierwszą osobą, która o tym pomyślała był Moritz Wagner, niemiecki badacz i historyk przyrody.
Wagner wcześnie rozpoczął karierę geografa i opublikował wiele książek geograficznych o Afryce Północnej, Bliskim Wschodzie i Ameryce Tropikalnej. Był również zapalonym przyrodnikiem i kolekcjonerem, i to właśnie z tej pracy jest najbardziej znany wśród biologów. Ernst Mayr, ewolucjonista i historyk biologii, opisał znaczenie Wagnera. str. 562-565
Podczas trzyletniego pobytu w Algierii Wagner zajmował się między innymi badaniem bezskrzydłych chrząszczy Pimelia i Melasoma. Każdy rodzaj dzieli się na kilka gatunków, z których każdy jest ograniczony do odcinka północnego wybrzeża pomiędzy rzekami, które schodzą z gór Atlas do Morza Śródziemnego. Gdy tylko przekroczy się rzekę, pojawia się inny, ale blisko spokrewniony gatunek.
"... początkowy gatunek może powstać tylko wtedy, gdy kilka osobników przekroczy granice swojego zasięgu... powstanie nowej rasy nigdy się nie powiedzie... bez długiego i ciągłego oddzielania się kolonistów od innych członków ich gatunku".
Był to wczesny opis procesu jednego z rodzajów specjacji geograficznej. W 1942 roku został on ponownie wprowadzony przez Mayra, a znaczenie specjacji geograficznej stało się jedną z głównych idei syntezy ewolucyjnej.
Innym terminem określającym specjację geograficzną jest specjacja allopatryczna. Allopatria oznacza "inny teren".
Wolne wyspy
Wyspy wulkaniczne powstają bez życia, a całe życie musi przybyć niesione przez wiatr lub wodę. Wiemy z wysp hawajskich i z Wysp Galapagos, że wszystkie formy życia zmieniają się, gdy docierają na wyspy z kontynentu.
Wyspy Hawajskie, zajmujące około 6500 m² (17000 km2), posiadają najbardziej zróżnicowaną kolekcję muchówek drosophilid na świecie, żyjących od lasów deszczowych po górskie łąki. Znanych jest około 800 hawajskich gatunków drosofilidów.
Badania wykazują wyraźny "przepływ" gatunków z wysp starszych na nowsze. Zdarzają się również przypadki kolonizacji z powrotem na starsze wyspy, a także pomijania wysp, ale są one znacznie rzadsze.
Według datowania radioaktywnego potasowo-argonowego, obecne wyspy pochodzą sprzed 0,4 miliona lat (mya) (Mauna Kea) do 10mya (Necker). Najstarszym członkiem archipelagu Hawajów wciąż znajdującym się nad morzem jest atol Kure, który może być datowany na 30 mya.
Sam archipelag, powstały w wyniku przesuwania się płyty pacyficznej nad gorącym punktem, istniał znacznie dłużej, co najmniej do kredy. Wyspy hawajskie oraz dawne wyspy, które obecnie znajdują się pod powierzchnią morza, tworzą łańcuch gór podwodnych Hawaje-Emperor, a wiele z tych podwodnych gór to guyoty.
Wszystkie rodzime gatunki drosofilów na Hawajach wywodzą się od jednego gatunku przodka, który skolonizował wyspy około 20 milionów lat temu. Późniejsza radiacja adaptacyjna została pobudzona przez brak konkurencji i szeroki wachlarz wolnych nisz. Chociaż możliwe jest skolonizowanie wyspy przez pojedynczą ciężarną samicę, bardziej prawdopodobne jest, że była to grupa tego samego gatunku.
Na archipelagu hawajskim są też inne zwierzęta i rośliny, które przeszły podobne, choć mniej spektakularne radiacje adaptacyjne.
Gatunki pierścieni
W biologii, gatunek pierścieniowy to połączona seria sąsiednich populacji, z których każda może krzyżować się z populacjami sąsiednimi. Dwa końce łańcucha zachodzą na siebie.
Dwie populacje końcowe w serii są zbyt daleko spokrewnione, aby mogły się krzyżować. Takie niehodowlane, choć genetycznie połączone populacje "końcowe" mogą współistnieć w tym samym regionie, zamykając w ten sposób "pierścień".
Gatunki pierścieniowe dostarczają ważnych dowodów na ewolucję: ilustrują, co dzieje się w czasie, gdy populacje genetycznie się różnią. Richard Dawkins zauważył, że gatunki pierścieniowe "pokazują nam tylko w wymiarze przestrzennym coś, co zawsze musi się dziać w wymiarze czasowym".
Trudno jest jednak znaleźć prosty, nieskomplikowany przykład.
Mewy Larus
Klasycznym przykładem gatunków pierścieniowych jest pierścień gatunków okołobiegunowych mew Larus. Zasięg tych mew tworzy pierścień wokół bieguna północnego, nad którym zwykle nie przelatują pojedyncze mewy.
Ogólnie rzecz biorąc, mewy mogą w pewnym stopniu krzyżować się ze swoimi sąsiadami, z wyjątkiem dwóch końców pierścienia.
Mewy czarnogrzbiete i mewy śledziowe różnią się od siebie na tyle, że nie dochodzi między nimi do hybrydyzacji; w związku z tym grupa mew tworzy kontinuum, z wyjątkiem miejsc, gdzie te dwie linie spotykają się w Europie.
Choć rzeczywistość jest o wiele bardziej skomplikowana, dobrze pokazuje ogólną ideę.
Salamandry Ensatina
Salamandra Ensatina jest gatunkiem pierścieniowym występującym w górach wokół kalifornijskiej Central Valley. Zespół ten tworzy wokół gór kształt podkowy. Choć między każdą z 19 populacji wokół podkowy może dochodzić do krzyżowania, podgatunek Ensatina eschscholtzii na zachodnim końcu podkowy nie może krzyżować się z Ensatina klauberi na wschodnim końcu. Jest to ilustracja "prawie wszystkich etapów procesu specjacji" (Dobzhansky). Richard Highton argumentował, że Ensatina to przypadek wielu gatunków, a nie kontinuum jednego gatunku.
Warkoczyk zielonawy
Wodniczka zielonawa (Phylloscopus trochiloides) ma kilka podgatunków, z których P. t . viridianus jest najbardziej znany w Europie. Jest to gatunek pierścieniowy, którego populacje rozchodzą się na wschód i zachód od Płaskowyżu Tybetańskiego, spotykając się później po stronie północnej. Ich relacje są dość zagmatwane.




Specjacja sympatryczna
Specjacja sympatryczna odnosi się do powstawania dwóch lub więcej gatunków potomnych z jednego gatunku przodka, zajmujących tę samą lokalizację geograficzną. Obecnie uważa się, że jest to bardzo częste zjawisko.
W przypadku specjacji sympatrycznej, gatunki różnią się od siebie podczas zamieszkiwania tego samego miejsca. Często przytaczanym przykładem specjacji sympatrycznej są owady, które stają się zależne od różnych roślin żywicielskich na tym samym obszarze.
Istnienie specjacji sympatrycznej jako mechanizmu specjacji było przedmiotem gorących sporów. Argumentowano, że dowody specjacji sympatrycznej są w rzeczywistości przykładami specjacji mikro-geograficznej. Ogólnie rzecz biorąc, obecnie nie jest to preferowane wyjaśnienie. Jest to raczej wynik hybrydyzacji pomiędzy blisko spokrewnionymi gatunkami, po której następuje dobór naturalny działający na potomstwo takich krzyżówek. Jednym z powszechnie akceptowanych przykładów specjacji sympatrycznej jest przypadek pielęgnic z jeziora Nabugabo w Afryce Wschodniej, który uważa się za wynik selekcji płciowej.
Speciation via polyploidization
Poliploidalność jest przyczyną wielu szybkich specjacji, ponieważ z potomstwa np. tetraploidalnych x diploidalnych kojarzeń często powstaje triploidalne, sterylne potomstwo.
Jednakże nie wszystkie poliploidy są reprodukcyjnie odizolowane od swoich roślin rodzicielskich, a przepływ genów może nadal mieć miejsce, na przykład poprzez kojarzenie triploidalnej hybrydy z diploidalną, w wyniku czego powstają tetraploidy.
Wiele z istniejących gatunków roślin i większość gatunków zwierząt przeszło w swej ewolucyjnej historii poliploidyzację. Rozmnażanie udanych gatunków poliploidalnych jest czasem bezpłciowe, przez partenogenezę. Z nieznanych powodów wiele organizmów bezpłciowych jest poliploidalnych.
Mucha głogowa
Jednym z przykładów ewolucji w pracy jest przypadek muchy głogowej, Rhagoletis pomonella, która wydaje się przechodzić sympatyczne rozmnażanie.
Różne populacje muchy głogowej żerują na różnych owocach. Odrębna populacja pojawiła się w Ameryce Północnej w XIX wieku, jakiś czas po wprowadzeniu jabłek, które są gatunkiem nierodzimym. Ta żywiąca się jabłkami populacja żywi się zwykle tylko jabłkami, a nie preferowanymi w przeszłości owocami głogu. Obecna populacja żywiąca się głogiem zwykle nie żywi się jabłkami.
Niektóre dowody wskazują na występowanie specjacji sympatrycznej. Muchy głogowe dojrzewają później w sezonie i potrzebują więcej czasu na dojrzewanie niż muchy jabłoniowe; ponadto istnieje niewiele dowodów na krzyżowanie się (badacze udokumentowali 4-6% wskaźnik hybrydyzacji).
Pojawienie się nowej muchy głogowej jest przykładem postępującej ewolucji.
Hybrydyzacja
Rzadko, gdy osobnicy różnych gatunków łączą się w pary, powstaje nowy gatunek. Zazwyczaj produkty takich hybryd są bezpłodne (nie płodne) lub względnie bezpłodne, a więc są eliminowane przez dobór naturalny.
Znaleziono jeden przykład nowego i udanego gatunku hybrydowego. Nowy gatunek jest mieszańcem wróbla włoskiego i hiszpańskiego, a podstawowe fakty zostały sprawdzone poprzez analizę sekwencji DNA z ich krwi. Ptak ten żyje we Włoszech w regionie, w którym żyją oba gatunki rodzicielskie. Nie rozmnaża się z wróblami hiszpańskimi, mimo że żyje z nimi obok siebie.
Sztuczna specjacja
Nowe gatunki zostały stworzone w wyniku hodowli zwierząt domowych, ale początkowe daty i metody nie są jasne. Na przykład owce domowe powstały w wyniku hybrydyzacji i nie dają już zdolnego do życia potomstwa z gatunkiem Ovis orientalis, od którego się wywodzą.
Bydło domowe, z drugiej strony, może być uważane za ten sam gatunek co kilka odmian dzikiego wołu, gaur, jak, itp.
Gatunki laboratoryjne
Najlepiej udokumentowane przypadki tworzenia nowych gatunków w laboratorium miały miejsce pod koniec lat 80-tych. William Rice i G.W. Salt wyhodowali muszki owocowe, Drosophila melanogaster, używając labiryntu z trzema różnymi wyborami siedlisk, takich jak jasne/ciemne i mokre/suche.
Każde pokolenie umieszczano w labiryncie, a grupy much, które wychodziły z dwóch z ośmiu wyjść, były od siebie oddzielane, by rozmnażać się ze sobą w swoich grupach. Po trzydziestu pięciu pokoleniach dwie grupy i ich potomstwo zostały odizolowane reprodukcyjnie z powodu ich silnych preferencji siedliskowych: łączyły się one tylko w obrębie preferowanych przez siebie obszarów, a więc nie łączyły się z muchami, które preferowały inne obszary. Historia takich prób opisana jest w Rice i Hostert (1993).
Diane Dodd była również w stanie pokazać, jak izolacja reprodukcyjna może rozwinąć się z preferencji kojarzenia w Drosophila pseudoobscura po zaledwie ośmiu pokoleniach przy użyciu różnych rodzajów żywności, skrobi i maltozy.
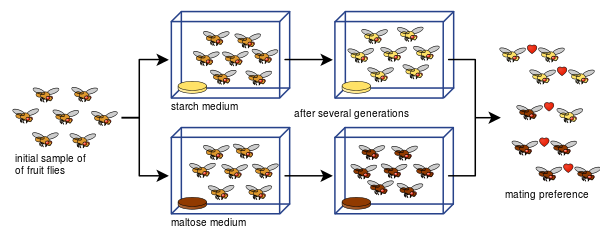
Eksperyment Dodda był łatwy do powtórzenia przez wielu innych, w tym z innymi rodzajami muszek owocowych i żywności.
Wzmocnienie (efekt Wallace'a)
Wzmocnienie jest procesem, w którym dobór naturalny zwiększa izolację reprodukcyjną.
Może to nastąpić, gdy dwie populacje tego samego gatunku zostaną rozdzielone, a następnie ponownie się zetkną. Jeśli ich izolacja reprodukcyjna była całkowita, to rozwinęły się one już w dwa odrębne, niezgodne gatunki.
Jeśli ich izolacja rozrodcza jest niepełna, to dalsze kojarzenie między populacjami wytworzy mieszańce, które mogą być płodne lub nie. Jeśli mieszańce są bezpłodne, lub płodne, ale mniej sprawne niż ich przodkowie, to nie będzie dalszej izolacji reprodukcyjnej i specjacja zasadniczo nastąpiła (np. jak u koni i osłów).
Powodem tego jest to, że jeśli rodzice hybrydowego potomstwa każdy z nich ma naturalnie wybrane cechy dla swoich własnych różnych środowiskach, hybrydowe potomstwo będzie nosić cechy z obu, i nie pasuje albo niszę tak dobrze, jak jeden z rodziców. Niska kondycja mieszańców spowodowałaby, że selekcja sprzyjałaby kojarzeniu asortatywnemu, co ograniczyłoby hybrydyzację.
Efekt ten nazywany jest czasem efektem Wallace'a od nazwiska biologa ewolucyjnego Alfreda Russela Wallace'a, który pod koniec XIX wieku zasugerował, że może on być ważnym czynnikiem w procesie specjacji.
Jeśli hybrydowe potomstwo jest bardziej sprawne niż ich przodkowie, to populacje połączą się z powrotem w ten sam gatunek na obszarze, z którym się stykają.
Wzmocnienie jest wymagane do rozdzielenia, gdy istnieje "strefa hybrydowa" pomiędzy dwoma formami gatunku. Strefy hybrydowe to regiony, gdzie rozbieżne populacje spotykają się i krzyżują. Potomstwo hybrydowe jest bardzo powszechne w tych regionach, które zazwyczaj powstają w wyniku wtórnego kontaktu rozbieżnych gatunków.
Bez wzmocnienia dwie populacje lub gatunki krzyżowałyby się w sposób niekontrolowany. Wzmocnienie może być wywołane w eksperymentach sztucznej selekcji, jak opisano powyżej.
Pytania i odpowiedzi
P: Co to jest specjacja?
O: Specjacja to proces powstawania gatunków. Jest to główna część biologii ewolucyjnej.
P: Co Darwin sądził o powstawaniu gatunków?
O: Darwin uważał, że większość gatunków pochodzi bezpośrednio od wcześniej istniejących gatunków, co nazwał anagenezą lub "ewolucją fitosocjologiczną".
P: Jak ogólnie postrzegano specjację w XX wieku?
O: W XX wieku naukowcy uważali, że większość gatunków powstała w wyniku podziału wcześniejszych gatunków, co nazywano kladogenezą. Uważano również, że podział ten był spowodowany lub wspomagany przez mechanizmy izolacyjne, takie jak separacja fizyczna.
P: Jak najnowsze badania zmieniły nasze rozumienie specjacji?
O: Ostatnie badania wykazały, że może dojść do hybrydyzacji pomiędzy spokrewnionymi gatunkami i że geny mogą być przekazywane poprzez te krzyżówki. Oznacza to, że izolacja reprodukcyjna nie zawsze jest konieczna do zdefiniowania gatunku, a allopatria (fizyczna separacja) może nie być wymagana do zaistnienia specjacji.
P: Co ma wspólnego separacja fizyczna z powstawaniem nowych gatunków?
O: Uważa się, że fizyczna separacja pomiędzy niegdyś współistniejącymi gatunkami jest głównym czynnikiem powstawania nowych gatunków, co ilustruje wiele przykładów omawianych w literaturze naukowej.
Powiązane artykuły
Autor
AlegsaOnline.com Powstawanie gatunków Leandro Alegsa
URL: https://pl.alegsaonline.com/art/92570
Źródła
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833